|
|
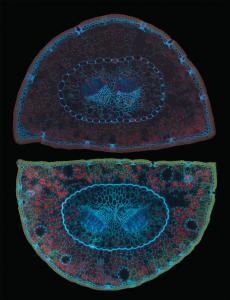
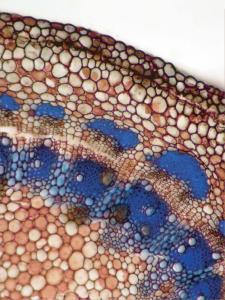
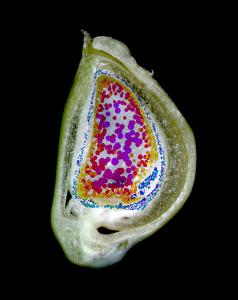
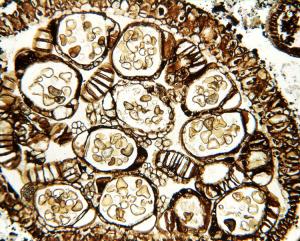
Plant Image CollectionMedia listVarious kind of media
|
















Copyright © 2024 The Botanical Society of America. 4344 Shaw Blvd., St. Louis, MO 63110 · Voice: 314-577-9566 · FAX: 314-558-9184 · Email: bsa-manager@botany.org

Items posted on the Botanical Society of America's website by the author/creator are licensed under a
Creative Commons Attribution-NonCommercial-ShareAlike 3.0 Unported License.
We value sharing, growing and learning together. In the spirit of fairness, we believe in the attribution of materials and ensuring the appropriate voices are in place when considering further use.